Dottorati di Ricerca

PhD Top Stories


PhD Top Stories
LA RIGIDITA� DEI TESSUTI INFLUISCE SUL METABOLISMO LIPIDICO TRAMITE SREBP1
Corso di Dottorato in Biomedicina molecolare
Un�equilibrata rigidit� dei tessuti � uno degli elementi necessari al corretto funzionamento degli organi. Tuttavia, � stato osservato che questa condizione � spesso compromessa in diverse patologie. Ad esempio, i tumori solidi sono pi� rigidi dei tessuti normali, caratteristica che contribuisce alla loro crescita e caratterizzazione maligna, suggerendo un legame fra la tensione dei tessuti e la formazione delle masse tumorali. Allo stesso modo altre patologie, come la fibrosi, sono caratterizzate da tessuti infiammati o danneggiati che, rispetto a quelli normali, presentano un elevato grado di rigidit�. L�eccessiva e continua stimolazione meccanica delle cellule in condizioni di rigidit� � capace di compromettere nel tempo il funzionamento di organi, come negli ultimi stadi di patologie del fegato, dei reni, dei polmoni e del cuore.
I meccanismi molecolari responsabili della rigidit� dei tessuti non sono ancora chiari, ma � sempre pi� evidente l�importanza del metabolismo nella reazione delle nostre cellule a sollecitazioni di natura meccanica. Comprendere quali siano le molecole e i processi chiave coinvolti nella risposta agli stimoli meccanici potrebbe avere dunque importanti ricadute sullo sviluppo di strategie terapeutiche per gravi patologie come fibrosi e cancro. In questo studio, � stata identificata SREBP1, una proteina nota per il ruolo chiave nella regolazione del metabolismo lipidico, come intermediario tra la risposta meccanica e la regolazione della biosintesi dei lipidi. � stato dimostrato che la regolazione meccanica esercitata da un�elevata rigidit� tissutale e dalla trasduzione del segnale meccanico da parte del citoscheletro di actina porta allo spegnimento della sintesi lipidica, sia con esperimenti in cellule di epitelio mammario umano, di fegato e polmone e in studi con animali di laboratorio come il moscerino della frutta Drosophila melanogaster (Fig.1)
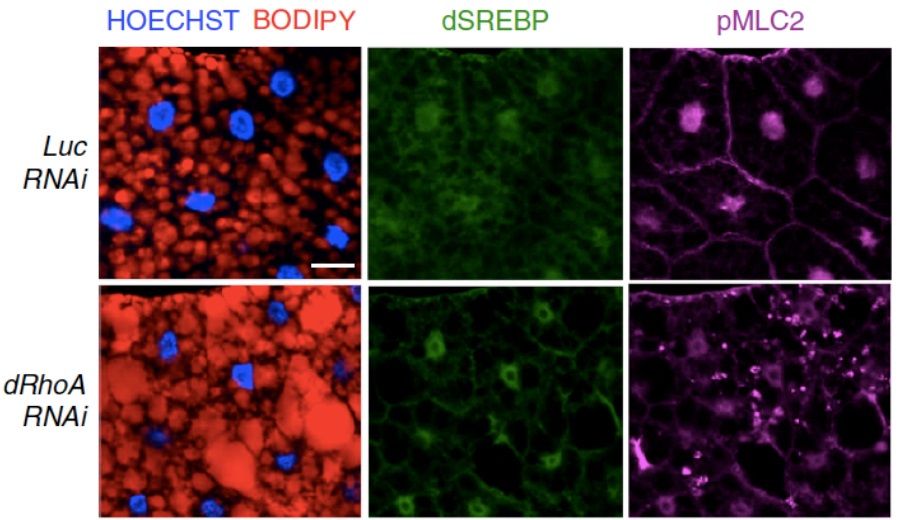
Figura 1: Immunofluorescenza di adipociti di larve di Drosophila in condizioni normali e di alterata segnalazione meccanica (dRhoAi) � in rosso: gocce lipidiche (BODIPY); in verde: SREBP1; in magenta: pMLC2- citoscheletro di actina; in blu: nuclei (HOECHST).
In risposta ad elevati stimoli meccanici, gran parte dell�energia prodotta dalla cellula viene utilizzata per il costante rimodellamento e contrazione del citoscheletro di actina, portando cos� la cellula a riprogrammare il suo metabolismo in modo da soddisfare le richieste energetiche di questi processi. � stato dimostrato che la proteina AMPK, che funziona come sensore del livello energetico della cellula, ha un ruolo importante nel mediare le vie di segnalazione delle giunzioni cellulari in risposta a stimoli meccanici, in modo di aumentare l�assorbimento di glucosio e, di conseguenza, la sintesi di energia, sotto forma di ATP. Il fatto che la biosintesi dei lipidi, che dipende da SREBP1, sia inibita dalla contrazione del citoscheletro, attraverso l�attivazione di AMPK, suggerisce che questo meccanismo potrebbe essere importante per integrare i segnali meccanici derivati dal microambiente con le richieste intracellulari, permettendo alla cellula di mantenere uno status energetico ottimale.
Il controllo esercitato dal citoscheletro di actina potrebbe essere un meccanismo fondamentale coinvolto nella regolazione del destino cellulare. Infatti, � gi� noto che gli stimoli meccanici sono strumentali per le cellule staminali per differenziare in osteociti (ambiente rigido) oppure in adipociti (ambiente lasso). Inoltre, � stato dimostrato che SREBP1 promuove l�adipogenesi, in quanto � un fattore importante nell�induzione del programma trascrizionale del differenziamento in adipociti. Quindi, in questo lavoro abbiamo scoperto che � in risposta a stimoli meccanici alterati che SREBP1 guida il destino cellulare delle cellule staminali mesenchimali in adipociti (Fig. 2)
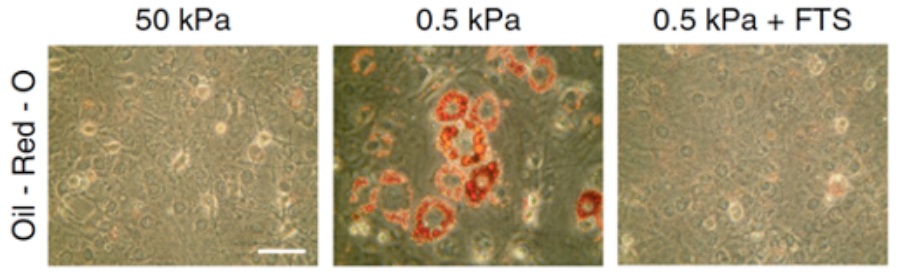
Figura 2: Differenziamento di cellule mesenchimali staminali in adipociti tramite colorazione con Oil-Red-O (gocce lipidiche). Le cellule sono state cresciute su matrici a diverso modulo elastico (50 kPa e 0.5 kPa) e trattate con uno specifico inibitore di SREBP1 (Fatostatina � FTS).
Inoltre, in questo progetto, � stato dimostrato che la contrattilit� dell�actomiosina regola la deposizione di grassi anche in Drosophila melanogater, attraverso l�attivazione di SREBP, sostenendo l�ipotesi che la coordinazione tra vie di segnalazione meccaniche e metaboliche pu� avere delle implicazioni a livello dell�intero organismo, come anche in condizioni fisiopatologiche nell�uomo. Infatti, in questo studio � stato dimostrato che in tessuti epiteliali mammari e polmonari, con un�alterata deposizione di matrice extracellulare e caratterizzati da un indurimento del microambiente circostante (in termini di densit� mammografica e fibrosi polmonare idiopatica), l�attivit� di SREBP1 � inibita, suggerendo che un�elevata stimolazione meccanica altera in qualche modo la corretta regolazione fisiologica di processi metabolici. La regolazione del metabolismo dei lipidi, esercitata dall�azione degli stimoli meccanici, potrebbe indicare un nuovo bersaglio terapeutico in queste patologie, ristabilendo le giuste propriet� della matrice extracellulare e, di conseguenza, una corretta omeostasi metabolica della cellula.
Autori ed affiliazioni
Rebecca Bertolio1/2, Francesco Napoletano1/2, Miguel Mano3, Sebastian Maurer-Stroh4/5, Marco Fantuz1/6, Alessandro Zannini1/2, Silvio Bicciato7, Giovanni Sorrentino1#, Giannino Del Sal1,2,82Dipartimento di Scienze della Vita, Universit� degli Studi di Trieste, Trieste, Italy.
3Center for Neuroscience and Cell Biology, University of Coimbra, Coimbra, Portugal.
4Bioinformatics Institute (BII), Agency for Science Technology and Research (A*STAR), 30 Biopolis Street, #07-01 Matrix, 138671, Singapore.
5Department of Biological Sciences (DBS), National University of Singapore (NUS), 14 Science Drive 4, 117543, Singapore.
6International School for Advanced Studies (SISSA), Trieste, Italy.
7Department of Life Sciences, University of Modena and Reggio Emilia, Modena, Italy.
8IFOM, the FIRC Institute of Molecular Oncology, Via Adamello, 16-20139, Milan, Italy.
#Current address: Laboratory of Metabolic Signaling, Institute of Bioengineering, Ecole Polytechnique F�d�rale de Lausanne, CH-1015 Lausanne, Switzerland.
Contatto
Rebecca Bertolio, email: rbertolio@units.itRiferimento bibliografico
Bertolio Rebecca, Napoletano Francesco, Mano Miguel, Maurer-Stroh Sebastian, Zannini Alessandro, Fantuz Marco, Bicciato Silvio, Sorrentino Giovanni and Del Sal Giannino.Sterol regulatory element binding protein 1 couples mechanical cues to lipid metabolism
Nature Communications 10, 1326 (2020)
DOI: 10.1038/s41467-019-09152-7